Taxa | Total abundance | %D | ||||||
| Dec | Jan | Feb | Mar | Apr | May | Total |
|
Osteichthyes: Atheriniformes |
|
|
|
| ||||
Odontesthes nigricans (Richardson, 1848) | 2 | 24 | 21 | 1 | 2 | 50 | 6.35 | |
Odontesthes smitti (Lahille, 1929) | 1 | 28 | 47 | 1 | 77 | 9.77 | ||
Osteichthyes: Carangiformes | ||||||||
Parona signata (Jenyns, 1841) | 1 | 1 | 0.13 | |||||
Osteichthyes: Clupeiformes | ||||||||
Sprattus fuegensis (Jenyns, 1842) | 4 | 4 | 0.51 | |||||
Osteichthyes: Perciformes | ||||||||
| Eleginops maclovinus (Cuvier, 1830) | 43 | 106 | 111 | 197 | 161 | 21 | 639 | 81.09 |
| Patagonotothen canina (Smitt, 1897) | 2 | 2 | 0.25 | |||||
Stromateus brasiliensis Fowler, 1906 | 1 | 3 | 4 | 0.51 | ||||
Osteichthyes: Salmoniformes | ||||||||
Salmo trutta Linnaeus, 1758 | 3 | 4 | 1 | 2 | 10 | 1.27 | ||
Chondrichthyes: Squaliformes | ||||||||
Squalus acanthias Linnaeus, 1758 | 1 | 1 | 0.13 | |||||
Total individuals caught | 49 | 163 | 187 | 201 | 167 | 21 | 788 | |
Total species caught | 4 | 5 | 7 | 4 | 4 | 1 | 9 |
|
ECOLOGICAL TRAITS OF THE PATAGONIAN BLENNY, ELEGINOPS MACLOVINUS (PISCES, ELEGINOPIDAE), IN THE RÍO GALLEGOS ESTUARY (SOUTHWESTERN ATLANTIC)
INTRODUCTION
Eleginops maclovinus is a euryhaline species, inhabiting coastal and estuarine environments, in temperate and sub-Antarctic waters from San Matías Gulf (along the Argentine Atlantic coast, 40°S) to Valparaíso (along the Chilean Pacific coast, 33°S), including the Malvinas (= Falkland) and Juan Fernández Islands (Guzmán & Campodónico 1973, Pequeño 1979, Brickle et al. 2005, Cousseau & Perrotta 2013). This species is characterised by a protandrous hermaphroditic cycle (Calvo et al. 1992, Brickle et al. 2005); and its growth rates showed marked differences between populations, ranging from 60 to 100 mm/year, which has been linked to differences in latitudinal distribution and food availability (Gosztonyi 1979, Licandeo et al. 2006). On the other hand, along the Patagonian coast, E. maclovinus is considered an essential component of the trophic web, as prey for several fishes, seabirds and marine mammals (Zama 1987, Brickle et al. 2005); while as a predator its feeding activity varies according to ontogenetic stage (i.e. juvenile and adult), habitat characteristics and food source (Guzmán & Campodónico 1973, Gosztonyi 1979, Pequeño 1979, Paves et al. 2005, Licandeo et al. 2006).
In recent decades, the study of fish species in coastal marine and estuarine environments of southern Atlantic Patagonia has increased (Caille et al. 1995, Torres et al. 2006, Martin & Bastida 2008, Torres 2009, Sánchez-Carnero et al. 2022). However, although these habitats are characterised by the presence of particular environmental conditions (e.g. large tidal amplitudes and extensive tidal flats) that favour the proliferation of a wide variety of species (Piccolo & Perillo 1999), community and ecological studies are still scarce. In addition, the Patagonian coastal area (in general) has experienced a significant increase in urban development and economic activities, and this growth could become a potential threat to the marine ecosystems of the region, causing irreversible changes in species diver- sity and abundance, as well as alterations in the trophic structure of these communities (Licandeo et al. 2006, Esteves 2007, Ferrari et al. 2007, Martin et al. 2019). In this sense, the study of fish populations and ecological traits are crucial for the management of exploited populations (Licandeo et al. 2006).
In southern Patagonia, small-scale artisanal fisheries are an important source of income for the local population (SERNAPESCA 2021, Sánchez-Carnero et al. 2022). Among the fishery resources, the Patagonian blenny Eleginops maclovinus is one of the main species caught, reaching ca. 20-200 tons per year (Caille et al. 1995, Licandeo 2006). Although this species has been well studied on the Atlantic and Pacific coasts of Patagonia (Gosztonyi 1979, Pequeño 1979, Zama & Cárdenas 1982, Isla & San Román 1995, Martin & Bastida 2008, Gastaldi & Gonzaléz 2009, Pequeño et al. 2010, among others), the fact that (1) a decrease in their abundance has been reported in some locations in northern Chile, related to the effects of pollutants on larval stages and overexploitation (Campos et al. 1998, Licandeo et al. 2006); (2) their contamination with antibiotics due to the consumption of pellets from salmon farming (Fortt & Buschmann 2007); (3) the modification of their biological traits related to climate change (Lattuca et al. 2017; Bilyk et al. 2018); (4) the increase of invasive species in Patagoniathat feed on E. maclovinus, such as the brown trout Salmo trutta (Zama 1987); and (5) their hermaphroditic life cycle, which makes them more sensitive to the selective pressure of fishing size than dioecious organisms (i.e. separate sex) (Licandeo et al. 2006); suggest that this species is highly vulnerable to anthropogenic activities. Thus, popula- tion studies of E. maclovinus provide complementary data on the status of the species at local and region- al scales, which is necessary for proper conservation of this species for fisheries (Licandeo et al. 2006). The aim of this work was to study some ecological traits of the Patagonian blenny E. maclovinus in an estuary of southern Patagonia during the artisanal fishing season, in order to (a) describe the fish fauna in the intertidal zone, (b) analyse differences in their monthly abundance and between tidal regimes (daytime and night tides), and (c) determine their feeding habits. This initial survey will provide important information for the design of further detailed studies, serving as a starting point for the development of natural resource conservation programmes.
MATERIAL AND METHODS
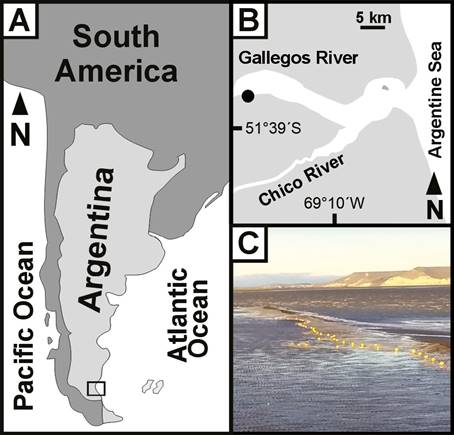
Study area: the study was conducted at the Río Gallegos estuary (51°35’S - 69°01´W; Fig. 1) on the southern end of continental Patagonia (Santa Cruz Province, Argentina). The estuary is located in the convergent mouths of the Gallegos and Chico rivers, which flow directly into the Atlantic Ocean (Piccolo & Perillo 1999). The macrotidal regime registers a mean amplitude between 2.8 to 11 m, with two high tides per day, in most cases one during daytime and the other at night. The southern coast of the estuary is dominated by extensive muddy intertidal flats, salt marshes and complex channels, while the northern coast is high with gravel beach plains (Perillo et al. 1996). The intertidal zone is characterised by the presence of a rich benthic community composed by molluscs, echinoderms, crustaceans, nematodes and polychaetes, which are the main food source for many fish and bird species (Torres 2009, Lizarralde et al. 2010, Ferrari et al. 2015, Martin et al. 2019, Pittaluga et al. 2019). The climate is cool (mean monthly temperature between-2 to 20 °C), dry (mean annual rainfall ca 250 mm) and windy (characterized by west winds blowing at an average speed of 35 km/h). Seawater temperature is influenced by the cold-temperate waters from the Malvinas current, va- rying between 4 to 13 °C (data obtained from CEADO-Argentina: http://www.hidro.gov.ar/ceado); while salinity va- lues range from 29.2 (in the inner part of the estuary) to 31.7 PSU (in the mouth), pH between 7.3 and 8.7, dissolved oxygen between 9.12 and 13.42 mg/l, and organic matter from sediments between 1.33 and 4.95 % (Pittaluga 2016).
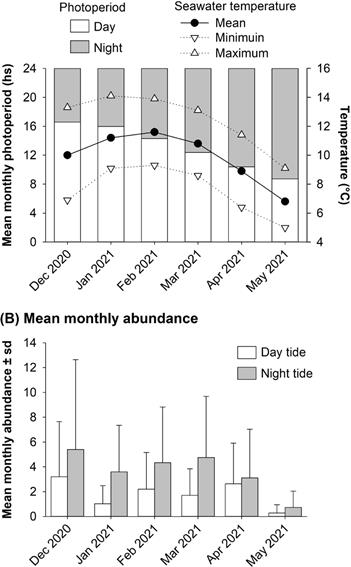
Field sampling, stomach contents and laboratory procedure: a field survey of fish was carried out 30 km from the mouth of the estuary, from 18/12/2020 to 21/05/2021, restricted to the artisanal fishing season, due to previous observations showed an occasional occurrence of E. maclovinus outside this period (Di Giácomo et al. 2015). Specimens were collected with a monofilament nylon gill net (length: 50 m, height: 3 m) with mesh sizes of 100 and 152 mm in the intertidal zone. The gill net was checked twice a day (i.e. daytime and night tides) during the low tide. Fish caught were counted and identified to species level (Torres 2009) and specimens of Eleginops maclovinus were stored in plastic bags for stomach content analy- sis. In the laboratory, each specimen was measured (total length). Stomachs of specimens from 12/01/2021 to 19/05/2021 were removed and their content analysed. Prey items were washed with running water and separated from stomach contents under a stereomicroscope. Items were fixed in ethanol 70 % and identified to the lowest taxonomic level possible using identification guides and specialised keys (Boschi et al. 1992, Forcelli 2000, Torres 2009, Häussermann & Försterra 2009). Monthly seawater temperature and photoperiod (day and night duration) were obtained from Servicio Meteoro- logico Nacional (http://www.smn.gob.ar) and CEADO-Argentina (http://www.hidro.gov.ar/ceado), respectively (Fig. 2A). Data analysis: for fish biodiversity analysis, the following data were calculated: total fish abundance (number of in- dividuals caught per month), percentage of dominance % D (percentage of individuals of a particular fish species in comparison to total individuals), total fish species caught (S´) . To determine if the Eleginops maclovinus catch va- ried between months and tides (daytime and night tide) a two-way ANOVA was performed (Zar 2009). A Student–Newman–Keuls (SNK) test was used when significant statistical differences in means were found (Zar 2009). Spearman correlation coefficient (r) was used to estimate the relationship between seawater temperature and photoperiod (day and night duration) with E. maclovinus specimens caught (Zar 2009). On the other hand, because stomach contents were highly digested and degraded, and some items were difficult to identify and count, prey abundance (% V) was eye estimated and calculated as the volume percentage occupied by each prey in comparison to the total volume of the stomach analyzed (Hyslop 1980, Baker et al. 2014). The frequency of occurrence (% FO) of prey items was monthly calculated as the percentage of total guts containing each item (Hyslop 1980, Baker et al. 2014). Statistical analysis was conducted in R statistical software (R Core Team 2022) using the package MASS and the level of significance was set to P = 0.05 (Venables & Ripley 2002).
RESULTS
Communitary and population analysis of Eleginops maclovinus
A total of 788 fish individuals were counted during the study period. The specimens collected included 9 identified species (Table I), reaching the highest values in February (7 spp.) and the lowest in May (1 sp.), remaining below 4 spp. during the rest of the months sampled. The patagonian blenny Eleginops maclovinus was the main species caught (ca. 81 % of total specimens collected), followed by the silversides Odontesthes smitti (ca. 10 %) and Odontesthes nigricans (ca. 6 %), the brown trout Salmo trutta (1 %), while fueguian sprat Sprattus fuegensis, butterfish Stromateus brasiliensis, cod icefish Patagonotothen canina, parona leatherjack Parona signata and spiny dogfish Squalus acanthias registered the lowest abundances (< 1 %). In addition, of the total fish caught on a monthly basis, E. maclovinus was the only species recorded throughout the entire study period and displayed the highest values (ca. 20-100 individuals per month), followed by O. nigricans and O. smitti (ca. 1-50 individuals per month), whereas the rest of species were observed occasionally (1-4 individuals per month).
E. maclovinus specimens showed a size range between 480-520 mm. The abundance of E. maclovianus showed a significant difference between months and tides (in both cases two-way ANOVA, P < 0.001; Table II, Fig. 2B). The mean values of E. maclovinus caught varied between 4 and 7 individuals per day from December to April, reaching the lowest values in May (1 individual per day; SNK test, P < 0.05). The comparison of tidal regimes established that E. maclovinus registered higher abundances during night (ca. 4 individuals per day) than daytime tide (2 individuals per day; SNK test, P < 0.05). Spearman correlation coefficient did not show a significant relationship between seawater temperature (r = 0.37; P > 0.05) and photoperiod (day duration vs daytime tide abundance, r = 0.49; night duration vs night tide abundance, r = -0.77; in both cases P > 0.05) with E. maclovinus specimens caught.
Source of variation | |||
Months | 5 | 4.44 | <0.001 |
Tides | 1 | 12.59 | <0.001 |
Months x Tides | 5 | 1.31 | 0.257 |
Taxa | Mean perecentage (%V) | |||||
Jan | Feb | Mar | Apr | May | Mean | |
| Mollusca: Bivalvia | ||||||
| Darina solenoides (King, 1832) | 73.65 | 93.39 | 82.91 | 70.46 | 58.33 | 75.75 |
| Crustacea: Peracarida | ||||||
| Acanthoserolis schythei (Lütken, 1858) | 0.17 | 1.86 | 2.97 | 1.00 | ||
| Edotia tuberculata Guérin-Méneville, 1843 | 0.77 | 0.76 | 0.16 | 0.34 | ||
| Mysida sp. | 4.17 | 0.83 | ||||
| Crustacea: Decapoda | ||||||
| Grimothea gregaria (Fabricius, 1793) | 8.85 | 4.24 | 11.74 | 2.75 | 5.51 | |
| Peisos petrunkevitchi Burkenroad, 1945 | 6.73 | 1.35 | ||||
| Osteichthyes | ||||||
| Odontesthes smitti (Lahille, 1929) | 0.38 | 0.12 | 0.52 | 0.20 | ||
| Patagonotothen canina (Smitt, 1897) | 1.92 | 0.23 | 2.97 | 4.17 | 1.86 | |
| Sprattus fuegensis (Jenyns, 1842) | 7.69 | 0.16 | 1.57 | |||
| Algae | ||||||
| Phaeophyceae sp. | 0.65 | 8.33 | 1.80 | |||
| Ulva sp. | 2.20 | 0.64 | 2.35 | 1.04 | ||
| Total items | 7 | 4 | 7 | 9 | 4 | 11 |
Feeding analysis of Eleginops maclovinus
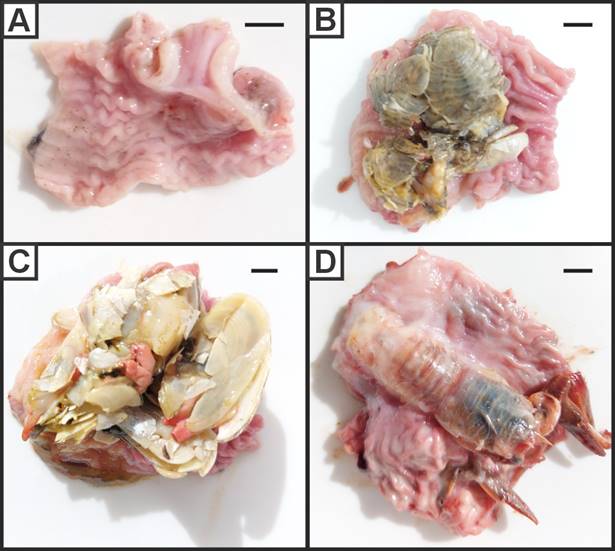
Feeding analysis revealed that from the total specimens of E. maclovinus (n = 448), 92.9 % had contents in the stomach while 7.1 % were empty (Fig. 3A). A total of 11 prey items were recorded (Table III), varying monthly between 3 to 9 spp. in January and April, respectively. The composition was represented by crustaceans (5 spp.), fish (3 spp.), algae (2 spp.) and molluscs (1 sp.). The most abundant species in the stomach contents were the bivalve Darina solenoides (ca. 76 %; Fig. 3B), followed by the decapod Grimothea gregaria (ca. 5 %; Fig. 3C). The peracarids Acanthoserolis schythei (Fig. 3D), Edotia tuberculata, Mysid sp., the decapod Peisos petrunkevitchi, the osteichtyes Odontesthes smitti, Patagonotothen canina and Sprattus fuegensis, and the algae Phaeophyceae sp. and Ulva sp., accounted for less than 2 % of the stomach contents in average.
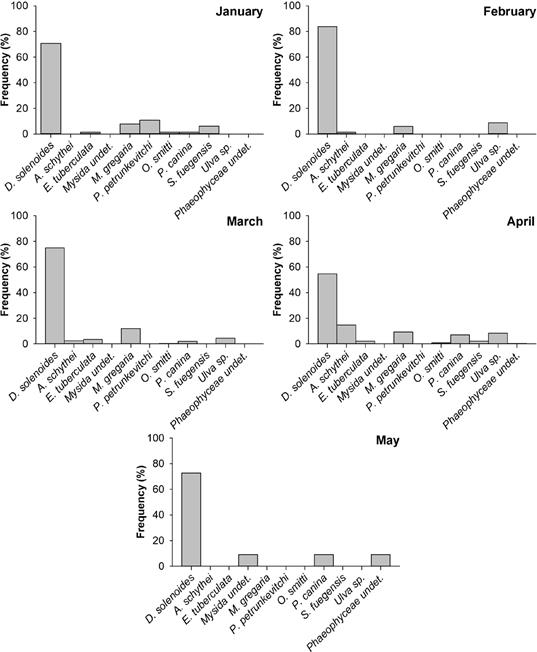
The taxon with the highest mean % FO in the stomach contents was D. solenoides (71.37 %), while the rest of species registered mean values lower than 7 %. Monthly % FO showed that D. solenoides was the dominant prey species with abundance percentage during all the months sampled, ranging between ca. 55 % (April) to ca. 83.82 % (March; Fig. 4); followed in January by P. petrunkevitchi (ca. 11 %), G. gregaria and S. fuegensis (ca. 6-8 %); in February by Ulva sp. (ca. 9 %) and G. gregaria (ca. 6 %); in March by G. gregaria (ca. 12 %) and Ulva sp. (ca. 9 %); in April by A. schythei (ca. 15 %), G. gregaria, Ulva sp. and P. canina (in all cases, ca. 7-9 %); and in May by P. canina, Phaeophyceae sp. and Mysida sp. (in all cases, ca. 9 %); while during the rest of the months sampled species registered a % FO lower than 5 %.
DISCUSSION
Previous studies have shown that Río Gallegos estuary shows a high species richness of fishes with a total of 35 species recorded (Caille et al. 1995, Torres et al. 2006, Torres 2009), of which 12 were classified as resident (i.e. species present throughout the year), 11 as seasonal (i.e. species recorded in a particular season) and 12 as occasional (i.e. species not recorded on consecutive occasions; Torres et al. 2006, Torres 2009). The present study found that from the total number of fish species in the estuary, only nine were registered in the intertidal zone during high tide, six of which were resident species (i.e., Odontesthes nigricans, Odontesthes smitti, Sprattus fuegensis, Eleginops maclovinus, Patagonotothen canina, and Squalus acanthias), while the remaining three were seasonal (i.e., Parona signata, Stromateus brasiliensis, and Salmo trutta). Resident fishes use the estuary as trophic and/ or breeding sites, explaining their highest abundance in the gill nets (Caille et al. 1995, Torres et al. 2006). However, some of these species were recorded occasionally (e.g., S. fuegensis, P. canina, and S. acanthias). This could be due to several factors, such as their distribution being restricted to the estuary mouth or the subtidal zone, their abundance is in decline or more gill nets are needed to increase the sampling effort (Torres 2009). On the other hand, the presence of seasonal species has been linked to their migratory and reproductive behavior. For example, species such as P. signata and S. brasiliensis extend their distribution from northern to southern latitudes in summer, affected by a warmer current (i.e. Brazilian current), entering in the estuary to feed (Torres 2009, Cousseau & Perrotta 2013). Meanwhile, species such as S. trutta move from the sea to the estuary for reproduction (Torres 2009, Cousseau & Perrotta 2013). However, to determine the population status of resident and temporal species in the estuary and explain the low abundances found in the present study, additional information is required. The monthly abundance variation of E. maclovinus may be attributed to its reproductive cycle.
Brickle et al. (2005) reported that the spawning period of this species from the Malvinas (= Falkland) Islands population occurs in deeper nearshore waters (> 40 m) during austral spring and secondarily in autumn, entering estuaries only to feed. Thus, the fact that adult individuals of E. maclovinus showed a total length close to the optimal size for sex-changing (360-500 mm; see Brickle et al. 2005, Licandeo et al. 2006), and that a high abundances of specimens were recorded from spring to summer in the inner part of the estuary (ca. 30 km from the mouth) supports the idea that the intrusion of E. maclovinusto the Río Gallegos estuary was mainly related to their foraging activity, obtaining nutrients and energy for their growth and gonadal maturation (Zama & Cárdenas 1982, Brickle et al. 2005, Licandeo et al. 2006, Di Giácomo et al. 2015). Reversely, the decrease in its abundance during autumn could be related to a migration process from shallow to deep coastal areas for reproduction.
Fish catchability may increase in response to environmental factors such as water temperature and photoperiod duration (Hammerschlag 2017; Leitão et al. 2018). However, no relationship was observed between these two parameters and the number of E. maclovinus caught. On the other hand, a significant difference in E. maclovinus abundance was recorded between daytime and night tides. It has been observed that fish species adopt different strategies to take advantage of light or dark conditions (Hammerschlag et al. 2017). Some species are more active at night to reduce their vulnerability to predators (Grol et al. 2008, Dupuch et al. 2009, Hammerschlag et al. 2010, 2017). Previous studies have shown that benthic macroinvertebrate abundances in the intertidal zone of Río Gallegos estuary is higher than in the subtidal zone (Pittaluga 2016, Lizarralde et al. 2017, 2019). Thus, the highest abundance of E. maclovinus during night tides could be due to this species moving from the subtidal to the intertidal zone to feed more safely. However, this hypothesis requires further evaluation, including factors such as differences in net visibility and moon phase effects (Hammerschlag et al. 2017). In general, the fish species collected during the study period mainly feed on plankton and fish (Torres 2009, Cousseau & Perrotta 2013). E. maclovinus is an exception as it has been classified as a predator of benthic species (Pequeño 1979, Licandeo et al. 2006, Martin & Bastida 2008, among others). The stomach contents of E. maclovinus from Rio Gallegos population also consisted solely of benthic organisms. The species composition was low (11 spp.) compared to other populations from coastal and estuarine areas of southern Patagonia (ca. 27-91 spp.) (Pequeño 1979, Isla & San Román 1995, Licandeo et al. 2006, Martin & Bastida 2008, Gastaldi & Gonzaléz 2009). Although, some populations from southern Pacific Patagonia registered similar values (ca. 11-14 spp.; Zama & Cárdenas 1982, Pavés et al. 2005, Pequeño et al. 2010). The low abundance of food items may be related to the lowest benthic species richness recorded in the Río Gallegos estuary (ca. 30 spp.), compared to nearby estuarine and marine populations, in which a highest abundance of food items and benthic species richness was recorded (32 and 69 spp. respectively; Martin & Bastida 2008, Lizarralde et al. 2017, Martin et al. 2019). However, it should be noted that studies where more food items were recorded, covered longer sampling periods (between 1-3 years; i.e. Pequeño 1979, Isla & San Román 1995, Licandeo et al. 2006, Martin & Bastida 2008, Gastaldi & Gonzaléz 2009). Therefore, it cannot be excluded that the lowest food items registered in Río Gallegos estuary could be related to the shorter sampling period, restricted only to the fishing season (ca. 6 months; Di Giácomo et al. 2015). Throughout the sampling period, the bivalve D. solenoides dominated the stomach contents of E. maclovinus in terms of both percentage volume and frequency of occurrence. These results suggest that D. solenoides plays a crucial role as an energy source for E. maclovinus, (Lizarralde & Pittaluga 2011, Lizarralde et al. 2018). D. solenoides is the dominant species in the middle intertidal level, with consistent density and biomass through the year (Lizarralde & Pittaluga 2011, Lizarralde et al. 2018, Gallardo et al. 2012, Martin et al. 2019). These results indicate that E. maclovinus was dependent on a specific prey, unlike other populations for which a wider prey item were found (in terms of quantity and occurrence) (Guzmán & Campodónico 1973, Pequeño 1979, Pavés et al. 2005, Licandeo et al. 2006, Martin & Bastida 2008, among others). Additionally, the presence of other food items in E. maclovinus specimens, such as crustaceans (i.e. Acanthoserolis schythei, Edotia tuberculata, Mysida sp., Grimothea gregaria, Peisos petrunkevitchi) and fish (Odontesthes smitti, Patagonotothen canina and Sprattus fuegensis), indicates that the feeding habits of E. maclovinus appears to be generalist and opportunistic, feeding mainly on those resources that are found in greatest abundance (Guzmán & Campodónico 1973, Pequeño 1979, Pavés et al. 2005, Licandeo et al. 2006, Martin & Bastida 2008).
In summary, this study concludes that E. maclovinus is the dominant species in the intertidal zone. The comparison of fish abundance between daytime and night tides suggests that this species is more active at night, possibly to avoid predation risk. The stomach contents revealed that E. maclovinus is an opportunistic species, feeding on most abundant and available benthic organisms. The present study will provide important information to design biological and ecological studies tending to determine the status of natural resources in the Rio Gallegos estuary, in order to develop appropriate legislation for the implementation of sustainable artisanal fisheries.
ACKNOWLEDGEMENTS .- This work was supported by the Agencia Nacional de Promoción de la Investigación, el Desar- rollo Tecnológico y la Innovación [grant number FONCyT PICT 2020-1855]; Universidad Nacional de la Patagonia Austral [grant number UNPA PI 29/A507]. The authors would like to thank the Universidad Nacional de la Patagonia Austral / Uni- dad Académica Río Gallegos (UNPA / UARG) and the Centro de Investigaciones y Transferencia de Santa Cruz (CIT Santa Cruz / CONICET). Thanks also to the reviewers for their comments and suggestions that largely improved the manuscript.
REFERENCES
Received on April 4, 2024
Accepted on October 16, 2024 Associate editor: T Changeux